南大洋的异养微生物受到低浓度有机碳(C)和铁(Fe)的双重限制。这些基本元素在细胞过程中紧密耦合;然而,在不同的环境条件下,原核生物对C和Fe的需求仍然缺乏研究。在这里,我们使用元转录组学和元蛋白质组学的组合来鉴定春季和夏末南大洋天然铁肥和高营养、低叶绿素水域中有机底物和铁的原核膜转运蛋白。
在两个位点,在转录本和蛋白质水平上,膜转运蛋白谱在季节之间都有明显的差异。当考虑特定的复合类时,这两种方法揭示了不同的模式。在转录水平上,季节性模式仅在属于每种转运体类别的基因亚群中观察到。在蛋白质水平上,有机化合物的膜转运蛋白在春季比夏季相对丰富,而铁转运蛋白则相反。这些观测结果表明,早春和夏末对有机碳和铁的需求增加。将转录本和蛋白质映射到50个宏基因组组装的基因组中,揭示了明显的分类群特异性季节性差异,指出了潜在的机会性进化枝,如假单胞菌科和氮菌科,以及表达转运蛋白更有限的类群,如Alphaproteobacteria和Flavobacteriaceae。
C和Fe膜转运体的联合研究表明,在不同的生产力制度下,微生物对这些元素的需求会发生季节性变化。不同形式的C和Fe的分类群特异性获取策略说明了不同微生物如何在不同季节在群落水平上塑造转录物和蛋白质表达谱。我们关于微生物类群C和Fe相关代谢能力的研究结果为它们在南大洋不同营养条件下C和Fe循环中的潜在作用提供了新的见解。
视频摘要
异养原核生物消耗了大约一半的初级产量,从而影响了通过海洋食物网的碳通量。原核生物对浮游植物衍生的有机物的转化决定了溶解有机物(DOM)的数量和质量,这些有机物可以在季节时间尺度上在地表水中积累,并最终通过翻转循环输出到深处[1,2,3]。
构成DOM库的各种单个化合物的浓度和组成决定了其总体生物利用度,从而决定了在不同时间尺度上可以转化的碳(C)的数量[4]。这一过程是由微生物代谢所必需的无机营养物质调节的。氮和磷是低营养海洋不同区域异养原核生物的主要生长限制因子[1],这些营养物可能导致表层水体DOM的季节性积累[2,5]。在南大洋,这些常量营养素在整个季节都保持高浓度,微量元素铁(Fe)是原核生物生长的限制或共同限制因素[5,6,7]。然而,这些先前的实验研究结果对南大洋原核生物对铁需求的时空变化提供了有限的信息。
海洋DOM由不同生物利用度的不同底物组成。随着分析方法的进步,DOM中所含有机化合物的复杂性变得更加容易了解[6,7]。同时,许多铁载体的化学表征和定量为铁生物地球化学提供了新的见解[8]。然而,各种原核生物分类群的有机底物和铁吸收策略仍然知之甚少。微生物转运体基因的表达谱可以作为描述有机物吸收模式的指标,并将其与分类学联系起来[9,10,11]。元蛋白质组学研究表明,广泛的有机分子参与微生物DOM通量[12,13,14]。亚转录组学为微生物对一系列高度不稳定的有机底物的吸收提供了新的视角,包括含氮化合物,如牛磺酸[15]和单碳化合物,如甲醇[16]、脂肪酸[15]和磺酸盐[17]。以类似的方式,不同的原核生物利用不同形式的无机和有机结合铁[18,19,20]。然而,对资源供应变化下的C和Fe转运体的联合研究仍然很少[21,22]。
本研究的目的是确定南大洋不同地点微生物群落中碳和铁化合物特异性转运体的季节性差异。我们的研究是在Kerguelen地区进行的,在该地区,天然铁施肥导致高营养-低叶绿素(HNLC)水域每年重复出现浮游植物大量繁殖[23]。这些花在不同的开花阶段深刻地影响异养原核生物的生长、活性和群落组成[24,25,26,27]。我们使用了元转录组学和元蛋白质组学相结合的方法,研究了早春和夏末两个具有不同季节生产力制度的地点的C和铁摄取模式,并进一步将基因表达数据映射到元基因组组装基因组(MAGs)中,以将系统发育与功能联系起来。
在初春(ES)(2016年10月6日至11月1日)和夏末(LS)(2018年2月18日至3月29日)的“南大洋与气候”(SOCLIM)巡航期间,采集了表层海水样本(10 m)。本研究选择了两个站点:一个站点位于高营养低叶绿素(HNLC)水域(KERFIX;50°40′S-68°25′E),其中一个站点位于Kerguelen中部高原上方的天然富铁水域(A3;50°38 ' S-72°02 ' E)(补充图1A)。在两次巡航中,除了KERFIX站在SOCLIM活动期间只采样一次(表1)外,两个站点都访问了两到三次。将Niskin瓶安装在CTD框架上进行海水采样,并使用酸洗管和60μm尼龙筛网将其转移到10-20 L聚碳酸酯瓶中。两次提取均采用相同的试剂盒和方法,所有重复的测序深度保持相同(补充材料;补充表1)。
表1简要说明研究地点的描述。2016年SOCLIM巡航期间进行了早春采样,2018年MOBYDICK巡航期间进行了夏末采样。所有数值均来自地表水(20米)一个早春和原核生物丰度(PA)数据来自Liu et al.(2019)[26],夏末来自Hernandez et al. (2021) [27]
采样和宏基因组分析,包括宏基因组组装基因组(MAGs)的构建,在其他地方进行了描述[21](补充材料;补充图2;补充表2、3和4)。简单地说,每个站点的6升海水使用47毫米过滤系统通过0.8微米膜过滤器(Isopore, Millipore)预过滤,并进一步在0.2微米Sterivex滤筒(Millipore)上收集。使用AllPrep DNA/RNA试剂盒(Qiagen, Hilden, Germany)从每个Sterivex过滤单元中提取DNA。按照制造商的指导方针进行DNA纯化,并在Agilent 2100生物分析仪/Agilent纳米DNA芯片(Agilent, Santa Clara, CA, USA)上检查DNA质量。Fasteris SA使用Illumina纳米文库制备试剂盒进行鸟枪文库制备,选择大小为550-bp。每个宏基因组在HiSeq 4000的一个完整通道上测序,每个宏基因组的成对末端reads为150-bp,每个宏基因组的reads在285 - 3.39亿之间。使用MEGAHIT v1.0.4[28]与默认参数和-预设的“meta-large”选项共同组装去污、裁剪和归一化的宏基因组序列,得到949228个至少1000 bp的片段(补充表2和3)。在宏基因组模式(-meta)下,使用Prodigal对开放阅读帧(orf)进行注释[29]。使用ConCOCT[30]、MaxBin v2.0[31]和metaBAT v2.0[32]这三种分类工具,利用metaWRAP流水线从组装的contigs中恢复个体基因组,共获得133个mag。每个MAG的完备性和冗余度通过CheckM进行估计[33]。133个mag的分类分类由GTDB- tk工具包[34]基于基因组分类数据库(GTDB v0.3.0)的classify_wf工作流确定,并通过基于单拷贝同源基因家族的系统发育推断进一步确认(补充表4)[21]。我们还将antiSMASH v5.1.2作为最近在海洋微生物组数据和转运蛋白上的应用[35,36],以识别我们的mag的生物合成簇,并在补品中添加了信息。表11和表12。
对于RNA提取,10-L海水通过0.8μm的膜过滤器(Isopore, Millipore)进行预过滤,使用142毫米的过滤系统(Geotech Equipment Inc.)和蠕动泵在0.2μm的膜(supportplus, Millipore)上收集细胞。RNA提取使用NucleoSpin?RNA Midi试剂盒(Macherey-Nagel, d
ren, Germany;补充材料)根据制造商的协议。使用Agilent 2100生物分析仪/Agilent RNA 6000纳米试剂盒(Agilent, Santa Clara, CA, USA)对提取的RNA进行定量和质量检查。测序前,使用RiboZero rrna -strand RNA协议对核糖体RNA进行酶处理,确保主要信使RNA测序,然后使用Illumina truseq -strand mRNA library Prep kit (Fasteris SA)构建cDNA文库。使用sortmeRNA和Fasteris使用的RiboZero rRNA耗尽试剂盒后,rRNA百分比在每个文库中总读取量的0.6至2.6%之间变化。所有元转录组(n=12)在HiSeq 4000的一条通道上测序,每个元转录组的150 bp配对末端reads产生2600万至3600万reads(补充表5)。将经过质量过滤的元转录组编码序列与元基因组共组装进行比对,并比较春季和夏季样本的计数表(补充材料;使用DESeq2 (v1.24.0)进行差异表达分析[37],鉴定出相对丰度变化显著的转录本(调整p值< 0.05)。此外,我们还使用ALDEx2[38]进行验证,采用了near等人[39]提出的第二差分丰度法,结果见附录。表13和表14。
对于蛋白质提取,20 l海水通过0.8μm的过滤膜(Isopore, Millipore)进行预过滤,使用142毫米的过滤系统(Geotech Equipment Inc.)和蠕动泵在0.2μm的过滤膜(supportplus, Millipore)上收集细胞。在两个季节进行两次过滤,并立即在- 80°C冷冻。使用Bayer等人[40]的改进方案从过滤器中提取全蛋白,提取的蛋白质进行变性聚丙烯酰胺凝胶电泳(SDS-PAGE),然后进行过夜胰蛋白酶凝胶消化(补充材料)。使用96孔板(SPEC 96孔C18, Agilent)提取和脱盐多肽,然后在2%乙腈/0.1%甲酸中重悬浮至0.2μgμL?1,然后注射到一维纳米流LC-MS /MS中(补充材料,补充表7)。使用归一化光谱丰度因子(NSAF)作为相对蛋白质丰度的代理,计算公式如下[41]:
其中,蛋白质k (PSM)中匹配肽的光谱计数总数除以蛋白质长度(L),然后除以所有N个蛋白质的PSM/L之和。
如上所述,Prodigal从组装的宏基因组contigs中共鉴定出了3003586个蛋白质编码基因。蛋白质序列通过CD-HIT-2D聚类[42,43](-c 0.9 -n 5 -d 0 - s2)来消除冗余。将得到的非冗余蛋白与Global Ocean Sampling (GOS)氨基酸序列数据库进行汇总,以便纳入更多来自海洋环境的蛋白质序列,这些蛋白质序列的读取长度较长,因此对于分析目的而言更加稳健[44]。另一轮cd-hit聚类(-c 1 -n 5 -d 0)去除相同的氨基酸序列,得到58,403,522条序列(补充图2)。为了鉴定个体宏基因组组装体中编码转运蛋白的基因,使用默认参数的eggnog-mapper v2[45],使用GhostKOALA[42]和eggNOG5.0[40,41]在KEGG数据库中查询预测的氨基酸序列。在KEGG数据库中分类为“转运体”的序列从元转录组和元蛋白质组中检索(作为电子补充材料提供)。转运蛋白家族和化合物特异性通过人工检查KO编号和转运蛋白分类数据库中的指定分类来验证[46,47,48,49]。为了确定具有MAG的蛋白质序列,将转运体对应的序列与高质量的规划宏基因组箱比对,并保留具有95%一致性和90%覆盖率的序列用于MAG特异性的进一步分析。为了特别关注碳和铁的利用,我们使用了现有的碳水化合物活性酶(CAZymes)的隐马尔可夫模型([50];http://www.cazy.org/)和铁特异性输运(补充表8和9)[51]。为了识别潜在的铁载体生产者,我们使用anti - smash (v5.1.2)[52]在MAG上进一步搜索BGC,相应的结果显示在补充表11和12中)。
摘要
背景
方法
结果与讨论
结论
数据和材料的可用性
参考文献
致谢
作者信息
道德声明
补充信息
相关的内容
搜索
导航
#####
自然施肥持续向克尔格伦高原以上的水域提供少量的铁,从而刺激了季节性初级生产和其他HNLC水域的相关食物网过程[23]。在目前的研究中,样本收集在每年初春的反复出现的浮游植物爆发在夏末post-bloom条件下(补充图1 b和C)。叶绿素a的浓度(背影)≈5倍高的高原站A3(1.54μg L?1)相比站KERFIX(0.32μg L?1)在HNLC水域在早春(表1)。这些差异更明显在夏末(≈0.39μg L?1 A3和≈0.17μg L?1 KERFIX)。两个地点的原核生物丰度呈现相反的季节模式,夏末的细胞丰度(A3处为8.94 × 108细胞L?1,KERFIX处为5.71 × 108细胞L?1)比早春(A3处为4.27 × 108细胞L?1,KERFIX处为2.89 × 108细胞L?1)高约2倍[26,27]。在克尔盖伦高原上方,我们观察到夏末溶解有机碳(DOC)浓度(54±1μmol L?1)略高于春季(52±1μmol L?1)。尽管不断输入铁,但不同地点之间的溶解铁浓度没有实质性差异,这一观察结果可以用地表水中铁的快速生物利用来解释[23]。在季节尺度上,溶解铁浓度在早春(A3和KERFIX分别为0.16 nM和0.13 nM)[53]高于夏季(A3和KERFIX分别为0.09 nM和0.07 nM)[54]。
meta -转录组学(MT)和meta -蛋白质组学(MP)分别鉴定了144和156个转运体转录本和蛋白质,其中98个转运体在两个数据集之间共享。转运蛋白的比例在3.5到12.3%之间,而它们在元蛋白质组中的比例在42.8到96.8%之间(补充图3)。属于atp结合盒转运蛋白家族(ABC)的膜转运蛋白的贡献最高(MT范围为2.8到10.8%;其次是外膜受体(OMR) (MT范围为0.22 ~ 0.77%;MP范围为3.6 - 19.6%)(补充图3)。之前在东南极洲的地表水[55]以及其他海洋区域和深层[12,56,57]中观测到这些转运体类型的类似分布模式。
在MT和MP数据集中,膜转运蛋白谱在不同季节之间存在显著差异(ANOSIM, p < 0.001)(图1)。我们进一步探索了MT和MP数据集的这些模式。编码铁转运相关蛋白家族的转录本在两个季节之间的相对丰度存在显著差异(图2)。
图1
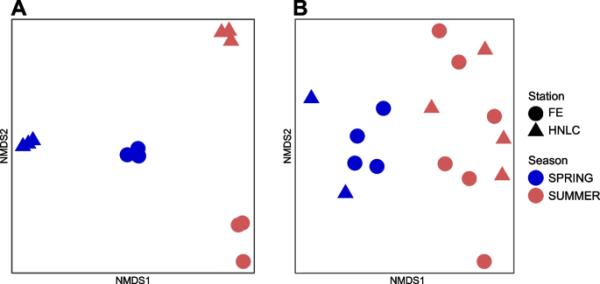
基于hellinger -transform Bray-Curtis不相似值的转运蛋白A和B元蛋白质组的非度量多维标度(NMDS)显示了每个站点和重访的生物重复(详见表S1)。两个季节的样本差异显著(亚转录组:1D胁迫=0.00001,ANOSIM, R=0.88, p < 0.001;元蛋白质组:二维应激=0.1;ANOSIM, R=0.84, p=0.001)
图2

KERFIX (A)和A3 (B)站点转运蛋白转录本丰度在社区水平上的季节性差异。每个点代表差异表达分析中可获得log2倍变化的转录本。负值表示转录本在夏末的丰度低于早春;正值表示转录本在夏末比早春更丰富
然而,大多数蛋白家族(与铁代谢转录调控相关的PF01325和PF02742除外)含有相对丰度增加或减少的转录本。因此,在MT数据中没有观察到铁相关转运蛋白的季节性趋势(图2)。同样,有机底物转运蛋白的比例在春季和夏季没有差异(补充图4)。
在蛋白质水平上进行了对比观察。大多数铁转运蛋白在夏季的相对丰度都高于春季,而有机C化合物转运蛋白的相对丰度则相反(图3)。
图3
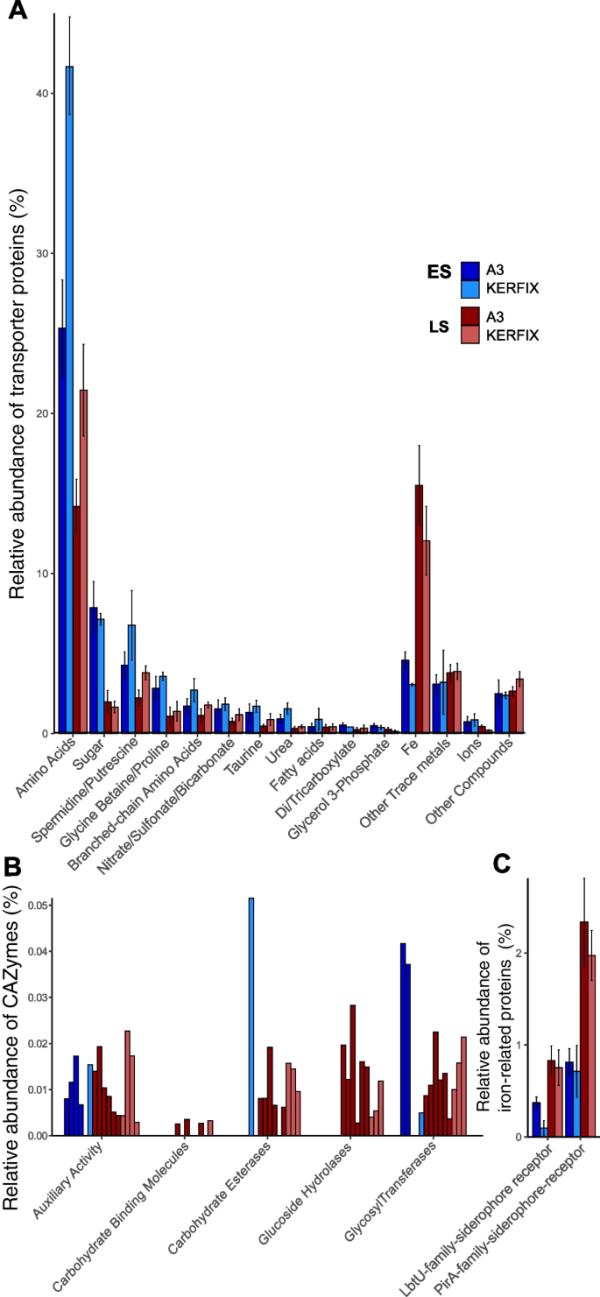
转运蛋白和糖活性酶在元蛋白质组中的相对丰度。柱状图显示了2个元蛋白质组的平均值,误差柱表示最小值和最大值(B除外,其中绘制了所有样本)。ES,早春;LS,夏末。A化合物特异性转运蛋白。B碳水化合物活性酶(CAZymes)。C铁铁载体转运蛋白(FeGenie分类)
含碳和含氮底物的转运蛋白最为丰富,且以氨基酸转运蛋白为主,分别占早春和夏末鉴定转运蛋白的25-42%和15-21%。亚精胺/腐胺转运体(春季为6.8%,夏季为2.2%)、甜菜碱/脯氨酸转运体(3.6%,夏季为1.1%)和支链氨基酸转运体(2.7%,夏季为1.1%)也出现了类似的季节性下降。
在这两个地点,牛磺酸、尿素、脂肪酸和硝酸盐/磺酸/碳酸氢盐的转运蛋白在两个季节的相对丰度都较低(< 2%)(图3A)。糖转运蛋白在春季占7.9%,在夏季占1.6%。辅助活性与cazy酶(如碳水化合物酯酶、葡萄糖苷水解酶和糖基转移酶)一起发生,在两个位点和季节都被检测到,占鉴定蛋白总数的0.05%(图3B)。然而,CAZymes在春季和夏末之间缺乏明显的差异。两个地点的铁转运蛋白(ABC和OMR)在夏末的相对丰度(占鉴定转运蛋白总数的15.5%)均高于早春(4.6%)(图3A)。同样,从春季到夏季,两种特定的铁载体受体LbTU和PirA的相对比例增加了2到4倍(图3C)。为了推断有机C和铁的摄取模式,我们计算了C转运体与铁转运体的比例之比(所有有机底物转运体与铁特异性转运体的总和)。该比值在春季显著高于夏末(高铁和低铁分别为22和10)(高铁和低铁分别为3和1)。
有机底物转运体和铁转运体在元蛋白质组中所占比例的明显季节性变化可能反映了早春和夏末对这些元素的原核需求发生了转变。这两个地点都观察到了这种时间变化,尽管施肥水域和不施肥水域的季节性生产力制度存在差异(补充图1BC)[24]。在早春,浮游植物活动的开始提供了生物不稳定的DOC,这是在之前非生产性冬季期间异养原核生物生长的主要限制[58]。清除多种化合物所必需的各自转运蛋白的投入[59]说明了原核生物对生物可利用有机质库变化的快速反应。
初春地表水中季节性高浓度的溶解铁和原核细胞中储存的铁都能满足一年中的这个时候原核生物的需要。在夏末,较高的原核生物丰度和代谢活动[27]表明c限制得到缓解。相反,夏季铁浓度低于春季,再矿化是铁输入的主要形式[60,61,62]。铁载体受体比例的增加(图3C)和转录调控(图2)表明有机结合铁的利用起着重要作用。铁载体的合成是能量昂贵的[63,64,65]。然而,当构成铁载体组成部分的碳和氮等营养物质相对过剩时(夏末可能就是这种情况),有机结合的铁可能成为铁的主要来源[66]。在此期间,具有生物合成和吸收铁载体结合铁的代谢能力的原核生物类群的生长可能更有利。
然而,当这些元素在夏末可能相对过剩时,有机结合的铁可能成为铁的主要来源,有利于具有代谢能力的原核生物类群,以生物合成和吸收铁载体结合的铁[18,19,20]。
结合mRNA和蛋白质组学的研究有望为细胞生理学提供深刻的见解,因为它们分别针对基因转录和翻译成蛋白质。然而,将这两种方法同时应用于环境原核生物群落的研究很少。在本研究中,我们观察到了亚转录组和-蛋白质组中季节性不同的转运蛋白谱(图1),但在特定化合物类别的水平上,这两种方法导致了不同的结果(图2和3,补充图4)。在解释这些结果时,应考虑固有特性和方法问题。原核mRNA的半衰期很短,变化很大(从1到46分钟不等[67]),mRNA的丰度比DNA少一个数量级,比蛋白质少四个数量级[68]。由于蛋白质在细菌细胞中存在的时间比编码它们的mRNA更长,因此需要考虑时间解耦[69,70,71,72]。此外,虽然蛋白质数据的半衰期平均较高,但两个实体也可能有持续的新生产,因此“常备库存”可能不受半衰期的影响。转录后和翻译后调控也是可以解释mRNA水平与蛋白质丰度不相关的过程[73,74],然而,我们认为这些可能起着次要作用。方法方面还包括与基于质谱的散弹枪蛋白质组学相比,RNA测序通常具有更高的通量和分辨率。
这些技术方面反映在我们的数据集中。膜转运蛋白在元蛋白质组中贡献了大部分蛋白质,但在元转录组中的相对贡献要低得多(补充图3)。对元转录组和元蛋白质组数据集的总体比较显示,功能注释基因的表达差异在定性上是一致的(补充图5和6)。然而,转录本的相对丰度与其相应蛋白质的相对丰度差异很大。正如早期研究发现的那样[59,69]。综上所述,本研究中观察到的不同模式可能反映了涉及蛋白质修饰和蛋白质水解的翻译后调控,可能导致特定蛋白质的积累。我们的数据表明,大量基因在两个季节都有差异表达,但这些基因转录本中只有一部分在元蛋白质组中被恢复,并且只能部分解释该方法的较高检测限。
然后,我们研究了转运蛋白在社区水平表达的季节性模式在个体分类群水平上可检测的程度。为了将有机底物和铁转运蛋白的表达与原核生物分类群联系起来,构建了宏基因组组装基因组(MAGs),并将从元转录组和元蛋白质组数据集中提取的转运蛋白编码基因序列映射到宏基因组箱中(完整性>=50%,冗余度<=10%;参见补充资料)。共获得了133个高质量的mag,涵盖11个门[21]。编码转运蛋白的基因转录本可以被定位到大多数MAGs(133个中的122个),而转运蛋白的蛋白质组肽可以被定位到50个MAGs(补充图7)。这可能是由于我们研究中使用的rna测序深度更高,与之前描述的基于质谱的肽分析相比,提供了更高的灵敏度。我们检索了两个数据集中存在的所有可以映射到MAGs的转运蛋白,并使用各自的转录本进行差异表达分析(补充材料)。
在比较春季和夏季群落时,观察到25个MAG中转运体的转录物丰度存在显著差异,并且这些季节变化是分类群特异性的(图4和补充图8)。在同一MAG中,除了属于根瘤菌属的一个MAG外,C或Fe转运体的丰度模式存在差异,但不是两种转运体类型都存在差异。转运蛋白编码转录本数量最多的是假单胞菌科和氮菌科,在季节间差异显著。其中,假单胞菌MAG 103的铁转运体的相对丰度在春夏季两个站点的相对丰度下降最为明显。相反,与之密切相关的假单胞菌MAG 62在夏末表现出相反的模式,两个地点的铁转运体丰度都较高(图4)。
图4
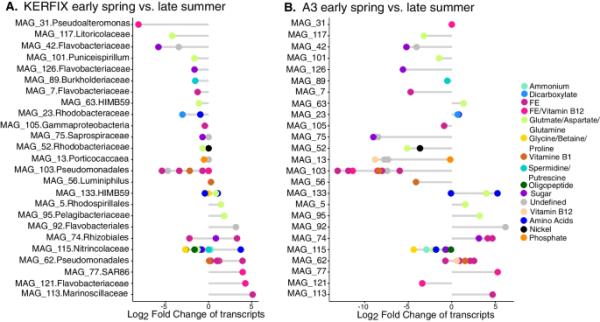
站KERFIX (A)和A3 (B)转运体转录本丰度的分类群特异性季节差异。负值表示夏末转录本丰度低于早春;正值表示转录本在夏末比早春更丰富。magid显示可识别的最低系统发育水平
在夏末,氮菌科MAG 115转录本中编码各种有机C底物(包括氨基酸)转运体的大部分转录本相对丰度较高。相比之下,分配给MAG 133 (Alphaproteobacteria HIMB59)的氨基酸转运体转录本在早春具有较高的相对丰度。这些分类群在春夏季铁转运蛋白(MAG 103和62)和有机C底物(MAG 115, MAG 133)表达中的作用在元蛋白质组数据集中得到了证实(图5)。膜转运蛋白的季节性表达视图说明了分类群中获取C和铁的不同生态策略,以及它们如何在群落水平上塑造表达谱(图2)。
图5
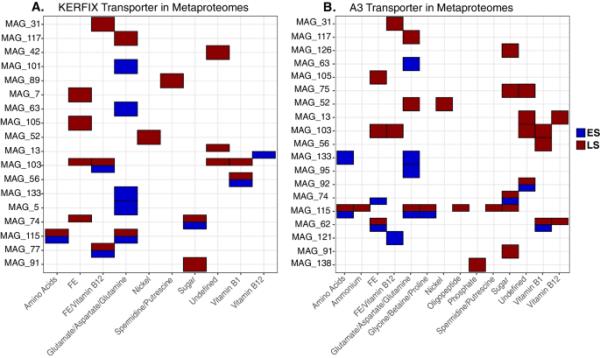
KERFIX (A)和A3 (B)的单个mag中不同化合物的转运蛋白的存在
我们的研究结果指向了潜在的机会性分支,如假单胞菌科和氮菌科[13,22],以及表达转运蛋白更有限的类群,如Alphaproteobacteria和Flavobacteriaceae。C-和Fe转运体的差异表达可能表明细胞需求和各自元素供应之间的时间解耦。在c -转运体的情况下,利用细菌铁蛋白中储存的铁的能力、低细胞铁配额或铁保留代谢可能是潜在的机制[20,75,76]。铁转运体的差异表达,而有机底物的差异表达保持不变,可能是由于细胞c代谢的适应[75,77]或独立于有机物供应的有效铁清除策略。这里观察到的早春和夏末之间的模式很可能也发生在更短的时间尺度上。
与此同时,我们的研究结果提供了南大洋两个不同站点在早春和夏末条件下的C和Fe转运体的分类群特异性模式,指向基因、转录物和蛋白质的复杂相互作用。由于每种方法的通量和分辨率不同,调和元转录组学和元蛋白质组学数据仍然是一项具有挑战性的工作。使用环境数据的解释使我们对化合物的供应和需求进行选择,这一点尚未得到澄清。最近的微流体研究已被用于利用宏基因组学在单细胞水平上阐明这些微调过程[78];然而,由于样品回收率低,这些目前不可能用于原位元蛋白质组学[78]。测量微生物细胞对各种化合物的原位吸收率仍然是海洋微生物组研究中的一个主要挑战。更好地了解这些分类群特异性特征及其在群落水平上的整合是未来的主要挑战,特别是如果我们想要了解微生物对变化环境的反应。
克格伦高原的一种测深法。站点KERFIX (1707m)和A3 (527m)的深度梯度位置。B. A3站近10年叶绿素a气候学和C. KERFIX站。绿线表示10年平均值;蓝线表示2018年(MOBYDICK巡航)。注意y轴上不同的刻度。
用于分析所有三个“组学”水平的生物信息学管道。A.宏基因组组装(个体组装和协同组装)和组合B.从宏基因组组装构建海洋蛋白数据库C. SortMeRNA提取mRNA后进行宏基因组定位D.利用宏基因组数据库进行宏蛋白质组学分析,并在eggNOG和KEGG中进行注释
运输家庭的相对比例。根据转运体数据库(TBDB)显示来自NSAF的A.元蛋白质组和基于总标准化转录本的B.元转录组不同转运体家族的相对比例的条形图。所有复件和三件均显示,并注明比例差异。ES -早春,LS -夏末,转运体类型以不同颜色表示:ABC - atp结合盒转运体复合物,AMT/NPP -铵通道转运体/硝酸盐,硝酸盐转运体,CUT -碳水化合物摄取转运体,OMP -外膜蛋白,OMR -外膜受体,SSS -溶质钠同向转运体,TRAP-T -三方atp非依赖性质周转运体,TTT -三方三羧酸转运体。
化合物特异性转运体转录本。ES:早春;夏末。转运体中化合物特异性转运体转录本的相对比例(见方法部分)。3个亚转录组的SD平均值。
共享KEGG转运蛋白热图。所有的亚转录组显示为三副本。标准化转录计数按行缩放,欧几里得按行和列聚类。元蛋白质组中蛋白质的存在是由它们在一个副本中的存在来定义的。转运体类型以不同颜色表示:ABC - atp结合盒转运复合物,AMT/NPP -铵通道转运体/硝酸盐,硝酸盐转运体,CUT -碳水化合物摄取转运体,OMP -外膜蛋白,OMR -外膜受体,SSS -溶质钠同向转运体,TRAP-T -三方atp非依赖性质周转运体,TTT -三方三羧酸转运体。
所有鉴定的KEGG转运蛋白热图。所有的亚转录组显示为三副本。规范化转录计数按行进行z缩放,按行和列进行欧几里得聚类。存在于元蛋白质组数据集中是指鉴定的蛋白质至少有一个重复。
50个元基因组的系统发育树,其中包含元转录组和元蛋白质组中转运蛋白表达的附加信息。由163个单拷贝基因计算得出的树。附加层代表MT-元转录组和MP -元蛋白质组中特定转运体类型的存在。
a站A3和b站KERFIX在早春和夏末转运蛋白的差异表达。左边的值代表早春的表达,右边的值代表夏末转运体类型的表达。MAG id显示可识别的最低系统发育水平。
补充材料。
抽样和复制描述。ES -早春,LS -夏末。补充表2。元转录组库的详细信息。补充表3每个元转录组库到协同装配的映射结果的详细信息。补充表4。用于宏基因组组装的库信息。补充表5。联合大会的成果。补充表6。133个元基因组组装基因组的GTDB分类描述。补充表7。元蛋白质组质谱分析结果。补充表8CAZymes HMM分析的元蛋白质组结果的相对标准化丰度。补充表9。FeGenie HMM分析的元蛋白质组结果的相对标准化丰度。补充表10DESeq2结果。补充表11。MAG序列中生物合成基因簇(BGC)的结果。补充表12结果MAG组份生物合成基因簇-与铁载体相关。补充表13春季与夏季铁相关蛋白的DESeq2结果。补充表14春季与夏季比较的Aldex2结果。
下载原文档:https://link.springer.com/content/pdf/10.1186/s40168-023-01600-3.pdf